My wife and I live on five hectares, mostly wooded, in central Massachusetts where I cut firewood, tend the garden, read Robert Frost and other poets, and observe the land, the sky and the living things nearby, often with camera in hand. I retired from Assumption College in Worcester, MA, in 2018. I taught biology and environmental science, and oversaw the design and construction of a new science building. I grew up outside of Chicago, the son of a teacher and a statistician. I started to pay close attention to the natural environment as an undergraduate, and continued on to graduate work in ecology. My fieldwork on plant-insect interactions has taken me to New York, California, Arizona, Massachusetts, New Jersey and Pennsylvania. My latest book explores the intersection of poetry and natural history: Stopping by Woods: Robert Frost as New England Naturalist. Watch this site for pictures and posts about the pond at the end of my road, plus thoughts about my life.
Tree nine is a white oak in Rutland, Massachusetts. I gave it that number when I was tagging trees that had lost a noticeable number of branches in an ice storm in 2008. Tree nine lost about half of its branches, but its neighboring trees lost nearly all of theirs. As a result, tree nine ended up growing less in the four years after the storm as it had in the three years before the storm, but not much worse than five or six years earlier. More sunlight compensated, at least somewhat, for having many fewer leaves. Other trees fared much worse.
Tree nine has been alive for a century, producing its thickest growth rings early in its life, but its growth declined steadily throughout its life. What happened?
The answer is simple. Nothing. Under constant, ideal conditions, any tree would have thick growth rings at first, with a steady decline in ring thickness throughout their lives. That’s the natural geometry of tree growth, as reflected in growth rings. Each year, a bigger tree makes more wood, but each year, it has to distribute that wood just inside the surface of a larger body (yes, the tree is a body). It might not be obvious how that will affect the width of each ring, year to year, but the result, both empirical and theoretical, is a steady decline in ring width each year (if you care, the decline follows a negative exponential curve).
So tree nine, in general, followed an expected pattern of growth. But there was also a whole lot of variation. Conditions were not constant, often not ideal, and sometimes destructive. Life in the forest is hard. Yet tree nine persevered, and does so to this day.
But the damage from 2008 didn’t heal completely, and eventually the tree was weakened by invading fungi. New wood couldn’t keep up with decaying wood, at least not in one of the major branches. In the summer of 2021, that branch broke, not completely, but more than enough to let the ends of the branch hit the ground, leaves and all.
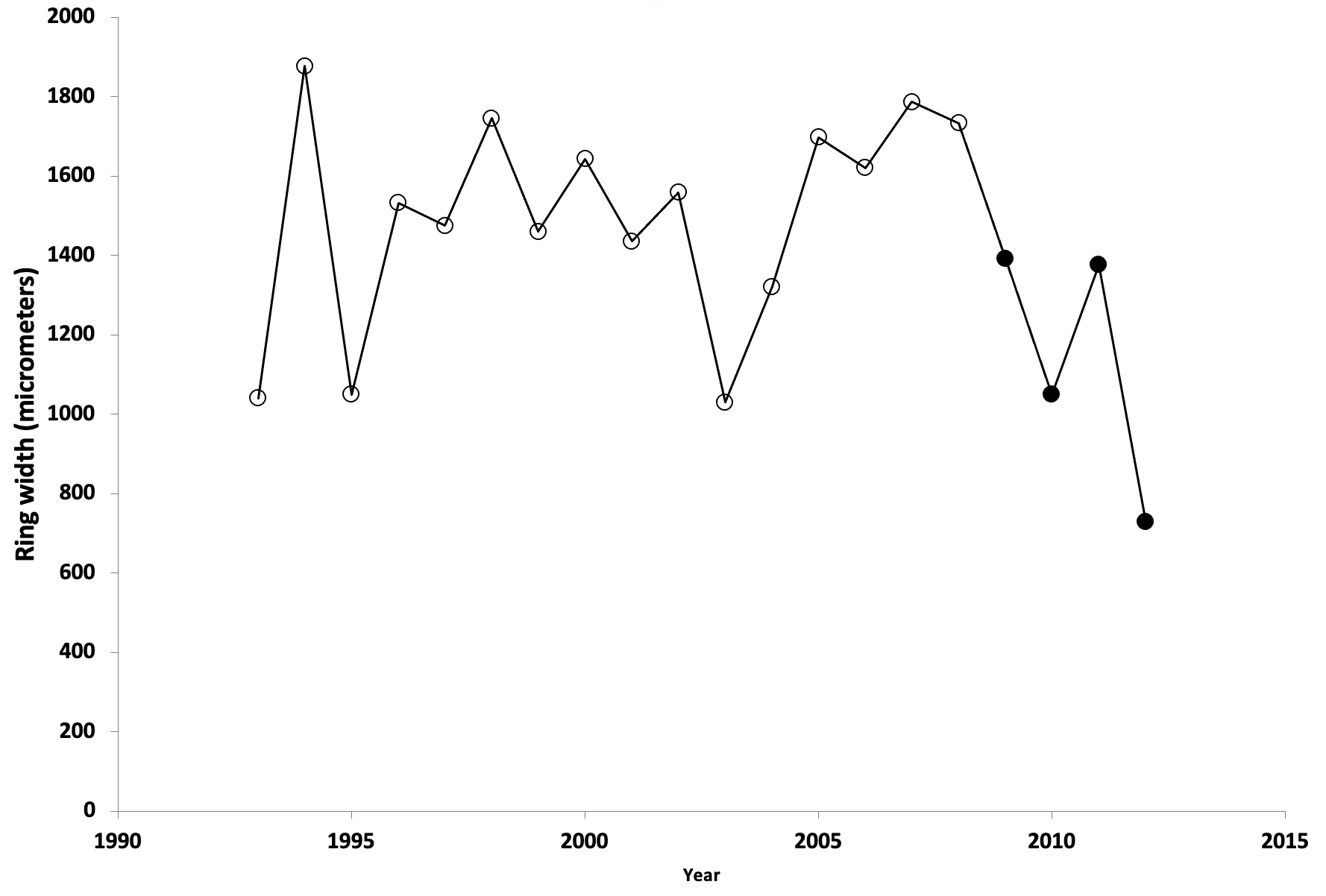
The base of the branch was still attached to the tree. There was enough good wood to retain a connection between trunk and branch, and good wood is tough stuff. This is what the growth rings look like in the fallen branch:
The bark is on the left side of this portion of the tree. The growth ring for 2021 ended just inside the bark, and began with the large cells a little to the right of the bark (the rings grow outward). The final ring is narrow because the branch broke off sometime in the spring or summer, and the branch stopped growing. The rings for three years after the ice storm (2008) are among the widest in this section of the fallen branch, but growth began to decline in 2012, probably because of the invasion of fungi that ultimately led to the failure of the branch in 2021.
The connection was not sufficient to sustain life in the fallen branch. The leaves of summer all died, and the stems and buds all died with them. The broken branch, with all the many branches stemming from it, is dead. I will harvest as much of the wood as I can reach to dry and then burn in my stove a year from now. I’ll leave the small twigs and branches and leaves for scavengers and decomposers. I will leave the parts out of reach for insects and woodpeckers.
Tree nine lives on, broken, but not dead. It still stands tall, and the surviving portions of the tree will grow. How long will it live? I have no idea, but I wouldn’t be surprised if it outlives me. Oaks, even wounded ones, can easily outlive people. Oaks give us hope.
Reference
Owen D. V. Sholes “Effects of ice storm damage on radial growth of Quercus spp.” The Journal of the Torrey Botanical Society 140(3), 364-368, (1 July 2013)
Goldenrods have finished blooming in central Massachusetts. They have also finished preparing for the future
The flowers produced seeds, one per flower, each with an embryo and a wind-catching pappus that could colonize a new location, maybe nearby, maybe far away. The seeds hang on to the parent plant, but the time it takes for them to leave varies enormously. Some break free quickly, and the last are still clinging to the old plant in the spring. Their variation probably increases the chances that at least some of them will be successful. If they all went at once, they might all get lucky – or they might all fail if there is some disaster that befalls the group. So spread them out over time and hope that some seeds will succeed. Of course, most will fail. That is the way with seeds.
The also vary genetically, most (all?) of the pollen having come from other plants, some nearby, some far away. These diverse offspring are mostly like their parents, but not identical. Will some grow better than their parents and leave lots of offspring? Maybe.
Goldenrods are perennial plants, sprouting up each year from tissues that have spent the winter or dry season underground. During the growing season, they produce new roots, and new rhizomes. The rhizomes are stems, botanically speaking, that spread underground. They don’t spread that far in any one year, and there might not be that many of them. But each year, if they succeed, they can take over a little more ground and make their clone a little more abundant in their habitat. Over time, they can take over a lot of land.
Some species of goldenrods from North America become invasive weeds when introduced into other continents. By seed and rhizome, they take hold and expand, just as they do in their original locations. They are among the many perennial weeds that have found ways to expand vigorously. Of course, there will be bad times and bad locations, and they will not take over the world. But they will keep trying.
I posted last year about bladderwort in a pond (Utricularia is the formal name – more on that in a bit). For several years, it bloomed like a carpet. In some years, it was a lush carpet, and in others, thinner, but still extensive.
This year, there were essentially no Utricularia in the pond. Probably not zero, but nothing even hinting at a carpet of flowers.
There are lots of animals that irrupt and become conspicuously abundant. Lymantria dispar, the soon-to-be-renamed gypsy moth, is one infamous example, and there are many others. Periodical cicadas are NOT an example – they just have a weird life cycle. But the annual cicadas in some places are an example. In sixteen years of field work among pinyon pines near Sunset Crater, Arizona, there were always a few cicadas around, except in one year when they were practically dripping from the trees. The next year, they were just around, not abundant. Why? I have no idea. And maybe nothing is more irruptive, and more mysteriously so, than the freshwater “jellyfish” Craspedacusta sowerbii. I saw it in a campus pond in 1979, and never again. It was spotted in 2020 in Walden Pond. It appears in a pond and then vanishes, only to appear in another pond years later. How does it do that? Who knows?
But are there irruptive plants? Algae blooms, sure. But vascular plants? The mast years of tree seeds or blooming years of desert annuals don’t qualify because they are always there, just not always reproducing or germinating. Perhaps gentians qualify, here this year, elsewhere the next, though they rarely get superabundant. I would not be surprised if there are truly irruptive plants, but I just don’t happen to know of any.
Except bladderwort. Utricularia just went through a boom and bust on my neighborhood pond, and I think that’s pretty amazing.
Now I have to admit that there’s a problem: I don’t know the species of bladderwort. It looks like the pictures of U. radiata (floating bladderwort) or U. inflata (swollen bladderwort), and I don’t have any specimens to check and confirm the identification. Both species have floating leaves and light yellow flowers. Both live in shallow ponds. But swollen bladderwort is said to be invasive, and invasive species often have the ability to grow quickly. That would be consistent with the sudden appearance of a carpet of plants on the pond. But their rapid demise was a surprise. So whatever species it is (I’m leaning toward U. inflata), I think it qualifies as irruptive.
So maybe – just maybe – the concern over an invasive bladderwort might be tempered a bit if their populations are likely to collapse after a few years. It would be helpful if we had more data, more observation of bladderwort populations over several years. If they consistently disappear, then maybe all we have to do is wait. That’s my hopeful hypothesis, and I look forward to learning whether I’m right.
In northeastern North America, there are a few species of goldenrod that conspicuously inhabit old farm fields, including Canada goldenrod, tall goldenrod, rough-leaved goldenrod, giant goldenrod, and early goldenrod. But there are many others. In my town, gray goldenrod (Solidago nemoralis), sharp-leaved goldenrod (Solidago arguta), and downy goldenrod (Solidago puberula) are three that are more common than one might think, but just not everywhere.
I’ve mentioned these species before, and I’ve even posted pictures of S. arguta earlier in the growing season, when it was just getting going. Sharp-leaved goldenrod is blooming now, but the only places I find it are along heavily-shaded dirt roads. Maybe that’s why some people call it forest goldenrod. The plants are tall, but the leaves are more widely placed on the stem than any species living in the open sun (except early goldenrod). My guess is that they don’t have the resources to make lots of leaves, so they spread them out in hopes of catching as many sun flecks as possible during the day. And they minimize one leaf shading another. They don’t produce a lot of flowers, but enough that you would notice them when you drive at a speed appropriate for a poorly-graded road.
Gray goldenrod is short, maybe half the height of the species that dominate old fields. I used six different fields in my research back in the 1970s, and in only one of them did gray goldenrod account for more than one percent of the plant cover. In Iowa, Patricia Warner and Robert Platt found gray goldenrod on the driest soils they studied. Massachusetts is not as dry as Iowa, but there are dry places here and there where the taller species might have trouble making a living. Those are places where gray goldenrod can maintain a foothold. One such place is in just a bit too steep to mow next to a field not far from one of our elementary schools. About 50 meters from a mesic slope covered with tall and Canada goldenrod, gray goldenrod is holding forth, blooming beautifully, with only grasses to cast shadows on their leaves.
Downy goldenrod used to be abundant behind our house when it was a field with small pine trees. Now those pines are several meters tall, casting too much shade for sun-loving goldenrods. Our neighbors sold off some timber about 25 years ago, and the open area by the road was another place for downy goldenrod, at least up until a few years ago. I checked last week and the birches have grown tall, too tall, it seems, for the goldenrods. I couldn’t find any.
But along the highway to the north, the west roadside has some downy goldenrods, and on the opposite side, with a sparsely-wooded wetland next to it, there is a thriving stand. They tend to be taller than gray goldenrod, but shorter than Canada goldenrod. They do best on nutrient-poor soil (reduced competition?), and many roadsides qualify for that distinction. Good to see some, compact and bright, finishing off the summer.
If a goldenrod looks a little different from what you expect, or is in a place that seems unusual, then it is probably not one of the more common species. These glimpses of something different are the first clue to the diversity of goldenrods. It’s a good time of year to appreciate how many kinds there are.
Most goldenrods have yellow flowers, but a few species of Solidago have white petals, and at least one of those (S. bicolor) has the common name silverrod. But perhaps we could call it platinum-rod, or palladium-rod, both metals having more of a white cast than silver. Whichever precious metal we use for the name, the flowers are showy. And the disk flowers are at least faintly yellow, like those in the rest of the genus (hence the species name bicolor?). In the riot of blooming at this time of year, a little variety is just fine.
Flowering plants produce seeds through the process of pollination, the release of pollen (containing sperm cells) from stamens, and the transfer of pollen to the receptive surface (stigma) of the pistil (containing egg cells).
Flowers
Goldenrod flowers (see my earlier posts) are clustered in flower heads. The flowers on the edge (ray flowers) have large petals and are solely female (no stamens). When a flower head opens, the first flowers to bloom are the ray flowers, which (presumably) provide nectar as a reward for any pollinating insect, and which can receive pollen.
The disk flowers in the middle of the head are both male and female. When the head first opens, the disk flowers are closed (see their blunt, closed ends in the photo above).
But later, the disk flowers open and the stamens thrust the anthers upward, exposing the pollen above the surface of the flower head (see photo below).
Insects
Many insects visit goldenrod flowers. I posted a one-minute video (Twitter and Facebook) of pollinator frenzy on a couple of goldenrod inflorescences. The video is full of wasps and bees, and goldenrods also attract beetles, ants, flies, butterflies, moths, and more. These insects get nectar and/or pollen as a reward (and some probably nibble on the tissues of the flowers). For some, the reward is immediate and short-term, fueling their activity during that day. For others, the reward is long-term, as they take the pollen and/or nectar back to their nest to share (for social insects) or to provision the nest for their young when they hatch.
Pollen is a durable, nutrient-rich food. Nectar is almost entirely sugar and water, an energy drink. A few species can evaporate the water to produce honey, a sugar solution so highly concentrated that it won’t spoil. I posted about honey earlier.
Pollination
As the insects cling to and walk on the flowers, they are almost certain to pick up some sticky pollen on their bodies. They might ignore the pollen, or try to clean it off, or eat some of it, or gather some of it to take with them, but no matter how carefully they groom themselves, some pollen grains are likely to be scattered on their bodies. When they move on to other flowers, some of those grains will rub off on stigmas, the topmost part of the female portion of a flower. If the pollen from plant A rubs off on a stigma of plant A, that will be a dead end. Goldenrods are self-incompatible (they can’t fertilize their own flowers).
But pollinators on goldenrods are greedy, abundant, and highly mobile. They will bump into each other, knock each other off of the flowers, and cause each other to fly to other flowers, some of which will be on another plant. Or they will drink the flowers dry and fly a bit to find more flowers, sometimes on another plant.
If the pollen from plant A ends up on the stigmas of plant B, then fertilization can get going.
Fertilization
The process of fertilization in flowering plants has several steps, and some of them seem at least somewhat improbable. But all flowering plants do it, and there are thousands of species of flowering plants. As complex as the process seems, they make it work.
When a pollen grain contacts the stigma of a flower, the grain “germinates.” A tube, called the pollen tube, grows from the pollen grain, and two sperm cells, both haploid, venture down the tube.
The stigma is the topmost portion of the pistil, the female portion of a flower, and the tube penetrates the stigma, the style below the stigma, and enters the ovary below the stigma. Inside the ovaries there are ovules, each surrounded by layers of cells, and each containing an embryo sac, a small structure with eight cells or nuclei, all of which are haploid.
The pollen tube finds its way into the ovary and then into an ovule and embryo sac. One of the sperm cells will fuse with one of the female haploid cells, the egg cell, and that fusion produces a diploid zygote, the first cell of a new plant. The zygote begins to divide and produces an embryo, which will stop growing and become dormant within the ovule. The ovule will eventually become a seed.
The other sperm cell inside the embryo sac fuses with two of the female haploid nuclei to produce a triploid cell (I warned you that the process was weird), and this triploid cell divides by mitosis multiple times to produce endosperm within the seed. Some plants have almost no endosperm (orchids), and others have a lot (coconuts). Goldenrods have enough to serve as a nutrition source for the seed when it germinates. Goldenrod seeds are small and light, easily blown around to places at some distance from the parent plant. But they also have a bit of endosperm to help the new plant to grow, wherever it lands.
Each goldenrod flower has one ovary with one ovule, so each flower produces, at most, one seed. The ovary provides a covering around the seed, which is technically a fruit, a dry fruit called an achene. As the ovary wall dries out, small filaments remain attached to the top of each seed. These filaments (the pappus) provide wind resistance and help the seed disperse away from the parent plant sometime in the next several months. Most of the seeds will fail for one reason or another, but there is always a chance that some will find a place to grow once the next spring rolls around.
Sexual reproduction
This whole sequence of events is sexual reproduction: flowers attract pollinators, pollen transfer leads to fertilization, fertilization leads to seeds and dry fruits, and dry fruits blow around in the wind. Each offspring has half of its genes from one parent and half of its genes from the other parent, and thus is genetically different from both them. Each unique seed then has a chance to colonize new habitats. It’s a risky business, but it has worked for an unfathomably long time.
If you feel lost in the details, take a look at the video on Twitter or Facebook. The frenetic pollinators can’t get enough of the goldenrod flowers. All that messing around leads to pollination, an essential part of producing another generation of goldenrods. And there it is, right by your house, along the road, or in whatever habitat your local goldenrods occupy. It won’t last forever, so take a look while it’s going on. If you miss it, it will all happen again next year. Nature is persistent.
Sometimes, a name tells you a lot. Shining flower beetles are shiny, they can be highly abundant on flowers, and they are, indeed, beetles. Some of their relatives are called shining mold beetles because they are abundant on fungi. All are in the family Phalacridae.
These beetles are also small, which probably makes the mold beetles easy to overlook, but the flower beetles are right there on top of the flowers, really hard to miss (if you stop to look). They are especially shiny in sunlight, and their brown color contrasts with the bright color of the flowers. They appear to be bite-sized for a bird, but I have never seen a bird grab one (though it would be so quick that the chances of seeing it are tiny, just like the beetles).
In the cool of the morning, the beetles are mostly between the flower heads, an indication that they spent the night there. In the middle of the day, they are often on top of the flowers, heads down, giving every indication that they are finding something on or in the flowers to eat. Their mouthparts probably cannot reach nectar, but pollen and tender floral tissue are right where their mouths are.
Grass-leaved Goldenrod (a necessary digression)
Decades ago, grass-leaved goldenrod was classified in the genus Solidago (S. graminifolia), but taxonomists decided that it, and similar species, should be in a separate genus, Euthamia. When it blooms, the flowers are yellow and look a whole lot like those of any other goldenrod around them.
So?
As you can see in the picture, shining flower beetles congregate on E. graminifolia flowers as soon as the first ones bloom. They spend little time on unopened flower buds, but can’t seem to get enough of the yellow blossoms. As far as the beetles are concerned, a goldenrod blossom is a goldenrod blossom, no matter how the plant is classified.
Where do shining flower beetles spend the winter? Good question. I would guess that the females lay eggs somewhere on, in, or near the plant, where the eggs remain dormant until the spring. The larvae live among flowers, but I don’t know whether the eggs wait to hatch until goldenrods are available. I suspect not, and I suspect that they have more than one generation a year. It would be interesting to find out.
Goldenrod plants are getting tall now, getting flowers ready to bloom, and becoming food for a variety of insects. One of the herbivores is conspicuous, if you take a little time to notice. Some stem tips look vigorous with leaves facing upward and outward and flower buds developing, like the picture above, but other stem tips are closed up and bent over, with the leaves facing inward and down. The outer surfaces are still green and, at a tissue level, seem healthy. But something is wrong.
Many plants show deformities, so these clusters of leaves might be some kind of developmental problem or pathogen infection. But no. If you pull the leaves apart carefully, there is white material holding the leaves together, and at the very center, there is a tiny moth larva. The white material is silk, spun by the caterpillar – yes, even one so small – tying the leaves together, holding them close to the caterpillar for protection and for food.
When I looked up “leaf tier” for goldenrods (pronounced “tie-er,” not “teer”), several species came up, representing multiple families, such as Tortricidae and Gelechiidae. I don’t have access to a microscope, so I can’t be sure which species I’m finding (and even with a microscope, species identification would be quite difficult), but they are doing well, whatever they are.
Most of the stem tips are undamaged, soldiering onward and upward. But they are beginning to show their age as insects find places on them and in them to live and to eat. The food web and growing season are marching forward together.
I already talked a bit about goldenrod hybrids and chromosomes. I will talk later about goldenrod reproductions, but briefly, they produce seeds that blow around (sexual reproduction) and rhizomes that spread underground (asexual reproduction). As a result of mutation, sexual reproduction, and hybridization, there is considerable genetic variation within any one species of goldenrod, and within any one population of a single species.
The photos at the top of this post are an example (all three are Solidago juncea, early goldenrod, July 30, 2021). They are growing on the same side of the same road, all within the same five-meter stretch of roadside. One is almost done blooming, one is in the middle of blooming, and one has yet to bloom. Yes, there could be some environmental differences between the locations of these plants, but it is highly unlikely that such minor differences would account from the considerable differences in phenology. No, the differences are mostly genetic, and are representative of the amount of genetic variation in goldenrods.
So what?
As it turns out, genetic variation has a measureable effect on what goes on in a field of goldenrods. Among the early goldenrod plants along my road, the differences might make it impossible for the earliest and the latest blooming plants to cross pollinate.
But there is more.
Gregory Crutsinger and his colleagues examined the significance of genetic diversity in an excellent field experiment published in 2006. First, they had to find a variety of goldenrods and clone them (they used tall goldenrod, S. altissima). Goldenrods are easy to clone because they spread vegetatively, but it takes time to grow enough clones. This experiment required long-term planning. And they didn’t merely assume that the different plants were genetically different. They compared the DNA of the different clones using a method called amplified fragment length polymorphism (AFLP). As expected, and confirmed, there were differences in the DNA between clones.
They planted the goldenrods in plots at a density of stems equivalent to those in many goldenrod fields in northeastern North America (12 stems per square meter). All plots had the same number of stems, but some plots had only one genotype, while others had three, or six, or twelve genotypes. They replicated each type of plot many times, and scattered each type of replicate throughout their test field to avoid bias by location. In short, they did it right.
They identified the arthropods (insects and arachnids) on the plants during the growing season, and measured the growth of the plants.
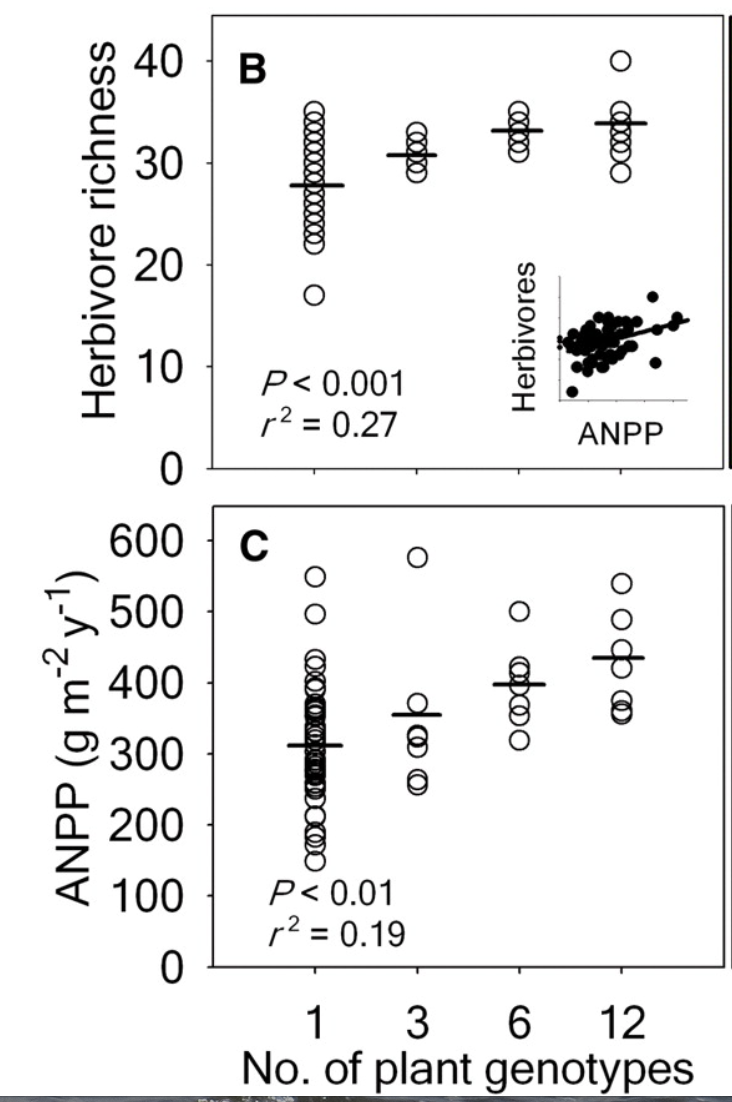
So what happened? The graphs show us what happened.
On average (the horizontal black bars among the circles), the greater the genetic diversity of the plants, the more species of herbivores, the more species of predators, and the more species of all arthropods were found in the plots (richness means the number of species). And the greater the genetic diversity, the greater the total growth of the plants in the plots (they measured growth as above-ground net primary production, ANPP).
The graphs show a lot of variation among the plots, and that the effects were statistically significant. Increased genetic diversity resulted in 20-35% increases in the average values of various measurements, which I would say is functionally significant.
I would not have predicted this much of an effect of the genetic differences among goldenrods of the same species, so I am delighted that someone else took the time to test it.
And I think it is highly likely that genetic variation within other plant species is also significant. Back in the 1970s, Janus Antonovics cloned a bunch of grasses that he collected in the wild, and checked to see whether the clones grew differently, whether alone or in competition with other clones. They did.
So genetic variation matters within a species.
When we talk about biodiversity (how many species in a habitat, region, or the planet), we need to include genetic diversity. Genetic diversity is the foundation for all diversity.
Reference
Gregory M. Crutsinger et al 2006. Plant genotypic diversity predicts community structure and governs an ecosystem process. Science 313: 966-968
You might have noticed the green sphere-like structures among some of the flowers in the pictures. Those are galls were produced by a species of gall midge (tiny flies). I hope to explore those in a future post.