My wife and I live on five hectares, mostly wooded, in central Massachusetts where I cut firewood, tend the garden, read Robert Frost and other poets, and observe the land, the sky and the living things nearby, often with camera in hand. I retired from Assumption College in Worcester, MA, in 2018. I taught biology and environmental science, and oversaw the design and construction of a new science building. I grew up outside of Chicago, the son of a teacher and a statistician. I started to pay close attention to the natural environment as an undergraduate, and continued on to graduate work in ecology. My fieldwork on plant-insect interactions has taken me to New York, California, Arizona, Massachusetts, New Jersey and Pennsylvania. My latest book explores the intersection of poetry and natural history: Stopping by Woods: Robert Frost as New England Naturalist. Watch this site for pictures and posts about the pond at the end of my road, plus thoughts about my life.
There is no pool in July. It is dead leaves and dust in a shallow depression. There are no understory plants, no young trees, just the shade of larger trees on the edge and in the high spots within the space. It is easy to walk through this dry spot, unlike the surrounding woods with young trees and fallen wood.
In autumn, it might get damp. But in winter, it fills with snow. One dry winter almost left it empty, but right at the end, a storm come through.
And then there is spring. It thaws, the water is full of dead leaves, it darkens like strong tea, and things begin to wake up. There are no leaves on the trees so the sunlight penetrates the murky water. Mosquitoes are common, as are beetles of several kinds, and the occasional water boatman. Phantom midge larvae bend and straighten in the brown water. Egg masses appear, frogs certainly, and maybe salamanders. I don’t obsess about the amphibians the way some people do, but these eggs are impossible to miss. They are colorful in their jackets of jelly.
But the headline attractions are fairy shrimp. They are orange and green and some other colors, with patches of iridescence. They are pretty big as adults (up to 2 cm), and slow, and they swim upside down. The males have weird appendages that resemble elephant trunks. Both males and females eat whatever they can find in the stew of organic debris, grow fast, mate, produce eggs, and die. It’s over quickly.
It has to be. The pond is drying. There is no outlet stream, so no fish can reach it. And no fish can live here when it dries up. The pond creatures would be gobbled up quickly by fish, so this is one of the few places they can succeed. But there is a price for safety, and that is time. There isn’t much time. Get your breeding done, and leave (if you’re a flying or crawling insect or a metamorphosed amphibian), or go dormant.
There are fairy shrimp eggs in that dust of July, in the soil and leaves, waiting. Summer is too soon. Autumn is too dry. Winter is too cold and frozen solid. Only spring will do. Once a year, spring is the season of explosion. Live fast, and go out in a blaze of glory.
When there are sharp seasons of the year, there will also be sharp rises and falls of growth, life, breeding, and death. If you can’t cope, get out. Come back when it’s better. And if you can cope, then congratulations on evolving a weird life cycle. Adaptation is alive and well in a vernal pond, and in every other thing around it.
In December, 2008, a storm came through the Worcester Hills in north-central Massachusetts with rain that froze on every exposed surface, and strong, gusty winds. Many trees lost limbs, some were snapped off entirely, or were bent horizontal to the ground. We lost power for a week.
Later that winter, I tagged dozens of oak trees on our property and estimated the amount of damage that they had sustained. Four years later, I took samples from each tree and measured the thickness of their growth rings. Tree five, a red oak, lost forty percent of its canopy, and though its annual growth before the storm often produced rings about one millimeter thick (occasionally much more), in the years right after the storm, the rings shrank to about half a millimeter. The tree was struggling.
In 2025, seventeen years after the storm, tree five stood out among its neighbors because of a substantial growth of bright orange fungi. It still had leaves, but it also had disease. That is, disease from the perspective of the tree. From the perspective of the fungus, life was very good indeed.
It had been a while since I had walked among the trees I measured after the ice storm. I recalled passing easily through the woods, with fewer low branches than decades before, a moderate accumulation of fallen branches and trees, and a fair number of small white pines whose flexible branches were easy to brush aside. Today, there were more fallen branches, and many of the small pines had dead branches, sometimes all the way up, and they were stiffly resistant to my passage.
What was the source of the fallen branches? Even the ice storm hadn’t left this profusion of limbs. What would be the fate of the pines, still deep in shade, and now with fewer needles to support their growth?
The impression I felt was one of decline, of trees large and small shedding the branches they could not support. In recent years, there had been dry spells, sometimes lengthy ones, during which at least one large pine had died, and more than one had dropped major branches (one of which crunched our small barn). And while most of the trees damaged by the ice were still doing well, a handful were dying or dead, and tree five was unable to defend itself against invading fungi. The ice demanded a price.
Things were changing. Things always change. But these changes, this decline, reminded me of a line from Robert Frost: “… what to make of a diminished thing.”
I don’t know what to make of it. I need more time. I need to look at what remains more carefully to see a larger picture.
At least as things were diminished, there were flames of fungi emerging from the base of the tree, this still-living tree, not going gentle into that good night.
This winter, I will walk again, and see what I can make of it in the snow.
This gorgeous green leaf glowing in the sunlight is pickerelweed. It lives on solar energy, incorporating carbon dioxide into sugar, and converting sugar into everything else. If it’s like a lot of other plants, about half of its sugar sustains metabolism, and the other half goes into growth and reproduction. Considering that it emerges from muck and lives in a shallow pond, it is surprisingly clean.
There is no shortage of water for the roots in the bottom of the pond (as long as it’s fresh water). The plants stay upright with strong cell walls, and with air in the stem that adds buoyancy. The air-filled stem also provides oxygen to the roots. Even before the stem reaches the surface in the spring, it is producing oxygen through photosynthesis in its green tissues, filling itself with bubbles and helping it rise upward to the light and keep the roots alive.
The plants produce flowers and seeds, all powered by the green stems and especially the leaves. And the leaves persist well into autumn, storing as much starch as they can in the roots, preparing to do this all again. Whether new plants from seeds, or persistent plants from roots, life differentiates itself from the non-living by being able to do it again, and again, and again, what Robert Frost called “The long bead chain of repeated birth.” So every year, the pickerelweed will rise to expose its beauty, and let us know that life continues to find a way.
The sun has risen, the waning moon is bright white against a blue sky with only a scattering of thin, pink, fluffy clouds, and the autumn colors are beginning to catch the rays of the sun. But as I walk along the road beside the trees, there is the distinct sound of drops – many drops, raindrops – hitting the leaves up in the canopy. It rained yesterday, but this morning, there are no rain clouds anywhere nearby. How can there be raindrops?
All last night, the leaves up in the canopy were fully exposed to the clear sky, open to the universe with nothing to hold in what little heat they might have absorbed beneath the clouds the day before. That heat radiated away, the temperature dropped below freezing, and the foliage was coated with ice (whether white or clear, I can’t tell). These are trees that can tolerate frost – they live in New England, after all. Their leaves are about to die anyway, and frost will not kill them any faster.
When sunlight hit the leaves after dawn, the leaves quickly warmed above freezing and their coating of frost melted rapidly, dripping down to the leaves below (amazingly little hit the ground, or me). And that’s what I heard, a false rain falling as frost released its grip. The melting will end soon, but for the moment, it is a delightful mismatch of sky and sound as this bright, sunny day begins with rain.
We were three weeks into fieldwork, and I had walked around campus to touch base with each group. They were pursuing projects of their own choice and their own design. Some were struggling, but still able to gather useful data. Some were realizing that they should have done something more, or something different, and now had to decide whether they had time to make the changes.
Then there was the vine project. All four of the women worked well together, and had a clear grasp of what they were doing. Were vines more abundant on trees in the sun than in the shade? Today, they had counted the vines (noting each species of vine) on the periphery of the stand of trees, and in what they defined as the interior. There were a lot more vines on the periphery.
I asked how they distinguished edge from interior, and they said it was kind of obvious to them. I asked how far in did the edge go, and how far in was the interior. They didn’t know, but they said that they could measure the distance to each tree and figure it out.
That was how things went with that group. They would make an observation or a count or a measurement, and share it. I would ask a few questions, and they would immediately figure out how to get more information to answer those questions, and other questions of their own. And they would go back to work. They knew (with my help) what all the trees and vines were, they knew where the trees were, how big they were, and which vines were found on which trees in which location. Were vines more abundant in sunny locations? Absolutely. Did tree size and tree species matter? Yes. Did the species of vines differ in their locations? It was hard to tell, but probably not much, if at all.
Later in the semester, they presented their work to the class, and wrote it up for me to read and grade, and it was all really good. The other groups did ok, but no group, that year, or any other year, did as well as this group. Some came close, but this group was always the best.
They graduated from college and went on to do a variety of things.
Not long after they graduated, I received an issue of the journal Ecology with an article about vines on trees in the southern Appalachians. Exposure to sunlight was a big deal for vines in that region, and I realized that all the work the vine group had done was sufficiently novel to be worthy of publication. I was able to contact all four women by email and ask whether they wanted to pursue publication. They all agreed, but they were busy with their new careers, and not experienced with formal writing for publication. So the task fell to me. Fair enough.
They sent me their notebooks, and I already had a copy of their report, so it didn’t take too long to produce a manuscript. I submitted it to a regional journal with them as first authors and me as last author. One of the reviewers was an author of the southern Appalachian study (she signed her helpful review). Our paper was accepted and published in 1998.
Ten years later, I applied for promotion. As a faculty member at an undergraduate college where teaching is our primary responsibility, I thought that I had accumulated enough publications, two of them with undergraduate coauthors, to qualify for an advancement in rank (as the policy document phrases it). I had done plenty of teaching and plenty of service, so I was reasonably confident that I had met the criteria in all three areas.
But there was a new Provost and a new President, both of whom thought they knew everything there was to know about faculty promotion. Their specialties were not in the natural sciences. Both of them had been Deans at universities before being hired for these new positions. Neither of them had attended, or been employed by, primarily undergraduate institutions. But now they were running our college.
So they decided that I did not deserve promotion because my publication record in particular was not up to snuff.
I asked the President for clarification. He told me to meet with the Provost. I asked to meet with the Provost. I heard nothing. Then, out of the blue, I heard that the President would, after all, meet with me. (I never saw that Provost again.)
In the meeting, the President acknowledged that my service met the standard for promotion. But he was unhappy about some negative comments on my course evaluations, so he concluded that my teaching did not meet the standard. I asked what the standard was for teaching. He repeated that I had too many negative comments.
Then he said that I had not published often enough to meet the standard for promotion. I asked how often faculty were expected to publish. More than I had, he replied.
But then he addressed his real concerns. I did not have enough sole-author publications (that’s how they did it in his field, after all). And publications with undergraduates “did not contribute to the discipline.” At that point, I realized that my appeal was utterly hopeless. He did not understand anything about research in the natural sciences, or about research at an undergraduate institution.
Four years later, under the third Provost to serve that same President, and with a few more small publications, I managed to convince them that I deserved promotion. Six years later, I retired.
But my promotion was less important than the education of the administration. My colleagues and I repeatedly emphasized the value of collaborative research and of engaging undergraduates in various research experiences. Undergraduate research is a worthy goal for faculty at a college, and a valuable experience for our students. Slowly, we convinced those in charge that working with undergraduates did, in fact, “contribute to the discipline.”
Those Provosts left the college years ago, and now, the President has left. We hope that their replacements will recognize and respect the scholarly work of faculty in all disciplines, and the engagement of undergraduate students in that work. It is especially rewarding for everyone involved when the students take ownership of the research. I’m glad that I was able to give students that opportunity while I was a professor.
Our paper:
Jennifer Buron, Danielle Lavigne, Kristine Grote, Rebecca Takis, Owen Sholes. 1998. Association of vines and trees in second-growth forest. Northeastern Naturalist 5: 359-362.
Our old science building, from the 1960s, had only one room that was easily available to students, and it sucked. The table was too big, leaving hardly any room for chairs. And the chairs were all rejects from elsewhere, and all were uncomfortable.
But there were always students in that room, doing whatever they had to do, conversing with one another, bursting out to find one or more faculty, whose offices were nearby, or to go to classes, which were also nearby. Proximity was a major reason why they used this horrible space. So was the simple act of getting together. Space mattered.
So when we designed our new science building (early 2000s), we wanted to include lots of student space, and the architects helped us get it.
There are expanded areas near landings and along railings, most of it close to classrooms and not far from laboratories. The soft chairs and small tables are used throughout the day.
We made clusters of faculty offices, and left a common area in each cluster with a table, white board, and simple, comfortable chairs. Some areas also have a microwave and mini-fridge, which students often ask to share, hardly ever abuse, and have never stolen.
There are also dedicated study rooms with tables and comfortable chairs. They have glass walls and lots of windows with nice views. While we were arranging the new furniture, the students helping us suggested that we add white boards. So we added white boards. We prevented faculty and administrators from reserving these rooms for meetings (the person in charge of reserving rooms was kind enough to take them off the list). Students always have priority. Students use these rooms all the time, write on the white boards all the time, and when they are preparing for exams, write on the glass walls, too. Yes, dry erase marker can be erased from glass.
One entrance to the building has a three-story atrium, and there are soft chairs, a few sofas, and small tables. Sometimes events require additional tables and chairs for a conference or meal, and students love to sit at the tables before and after the event for as long as the tables are present (no, they don’t mess up any tablecloths).
Another entrance has a two-story foyer and a small, carpeted area with a two chairs. These were the only wooden chairs we put in the building, and they are hardly ever used. We hadn’t expected them to be used because of the high traffic so close to the entrance, so we put in these exotic-looking chairs that were visually attractive, but uninviting. When someone brought a soft chair from another building, it was immediately used a lot. We should have put in soft chairs from the start.
Above that entrance is a balcony near three faculty offices and a conference room. We put in soft chairs and a small table, but students hardly ever sit there, and we still aren’t sure why. Maybe it’s because the conference room is often available, and has a white board. Maybe it’s because there are decent chairs in the nearby common area shared by the faculty offices. Maybe it’s too far from the classrooms, all of which were at the other end of the building. It’s a mystery.
The conference rooms are often used for faculty and administrative meetings, but there are plenty of times when they are empty. That’s when students move in. Students are particularly attentive to the presence of meetings in which food was served. When the meetings end, students grab the leftovers. But even without food, students know about and use the space.
One of the enclosed stairwells has windows, so we put in a nice wooden bench on one of the landings next to the window. In fifteen years, I have seen one student on that bench. I used it more often than the students. Maybe if the bench had cushions, it would have been more attractive. I certainly would have used it more myself.
Overall, even though we would change a few things about a few spaces, most of them worked as we had hoped. And there is no doubt that students would have used more spaces if they were near the action (faculty, classrooms, traffic) and attractive (soft seating and tables).
In short, student space matters.
And space doesn’t design itself. We consulted students during the design process, and faculty discussed it at length (Kim Schandel, Brian Niece, Stuart Cromarty, Ed Dix, and Steve Theroux all contributed). The architectural team at EYP, led by Heather Taylor, incorporated our wishes through their expertise, giving us the wonderful Richard and Janet Testa Science Center. Everyone deserves a huge amount of credit for contributing to a great process that led to a great result.
When should professors provide guidance to their students? We hand out the syllabus on the first day (or post it earlier) as a guide for the entire course. But is the first day the best time for all of that guidance? Some years ago, I stumbled upon an answer for that question: probably not. I learned this with regard to a semester-long project I assigned to my students.
Task
I taught introductory environmental science throughout my forty-year career. A few years in, I began asking the students to pick an article from the popular press (now, they get it from the web) and write a critique. But I wanted them to write an informed critique, so I split the task into three parts, due at different times, and allowed them revise the whole thing after they got my comments on all three parts.
Part one was a one-paragraph summary of the article, plus a list of the issues in the article that they would need to check. Part two was a compilation of information, from sources other than the article, about each issue they had identified (I called this the background section). Part three was the critique of the article, issue by issue and overall, comparing the article to what they had found from other sources (I called this the evaluation section).
Many (perhaps most) students had difficulty with one or more steps in the process. Some picked articles that were too short, or too long, or inappropriate for the course. Most had trouble identifying the issues. Some had trouble gathering and organizing information from other sources. Some had trouble using the information to evaluate the content and presentation of their chosen articles. In my comments, section by section, I tried to steer them in productive directions. I typed and printed my comments for each section so that they had a readable record of what they had done well, and of what they needed to revise. I always gave them hard copies (old habits die hard), but electronic transmission would obviously work instead.
When students had problems, I also referred them to the handout I gave them on day one about the task. Some found the handout useful, some didn’t, and some didn’t even have the handout when they needed it. I know this because they were supposed to keep everything in a folder and hand in the folder when each part of the paper was due. Most of them kept the handouts, and my comments, but some didn’t (using printed materials allowed me to provide replacement copies, when needed).
Timing
One semester, on the day I handed back the last set of comments on their evaluations, I also gave them a one-page schematic, based on the original two-page handout, that listed each part of the paper, what should be in it, and how it should be organized. I told them that they could revise everything, and that the schematic was a guide for their revisions. The classroom buzzed with comments, their voices rising and falling with the sounds of revelation. The most common sounds I heard were, “Oh, yeah!” They had been through the whole process of the critique once, and now they had directions for how to fix the problems. The students immediately began asking questions, each of them about a specific part of the critique as it applied to their particular article. Because I had read all their chosen articles and the drafts of their critiques, I could nearly always give them answers specific to their articles (the class ranged in size from 20-40 students).
Here’s my point: When the students got the guidance they needed when they needed it, the guidance was helpful. They were about to make the revisions that would earn them an actual grade, not just more comments. Now, things were serious. And now (not three months earlier) they could see how to get to a successful conclusion.
The original handout was also helpful, but it had not been particularly helpful on day one because it came too soon, before they knew the problems that they would actually encounter. Immediacy seemed to matter.
I was not always able to time my advice as well as I did for theses critiques, but I certainly tried, as did many of my faculty colleagues. We often exchanged ideas about how to provide various types of guidance. Sometimes, all we did was give students the same handout again, or reminded them of it, pointing out how the handout provided the information they should follow. Sometimes, we gave them a new, shorter handout, emphasizing essential aspects of the task. Sometimes, we would hand out or refer to the syllabus again, in whole or in part, to reinforce what the course was trying to accomplish. Often, we were able to provide the timely guidance that at least some students seem to need.
The moth Lymantria dispar has been feeding on trees in parts of North America since its introduction into the continent from Eurasia in 1869. By “feeding” I mean, “stripping every freaking leaf off” when the caterpillars are at peak abundance. In most years, the larvae are nearly impossible to find. But every so often (the frequency of outbreaks varies widely), there is a hoard of these hairy, irritating, munching critters, and trees in summer can be as bare as they are in winter. Where I live in central Massachusetts, we had an outbreak in 1981, and it was unpleasant to see the trees defoliated, to listen to the rain of droppings, and to have hoards of caterpillars all over the outside of the house. But we have had no outbreaks since, though pockets of abundance have appeared just a few miles away. We don’t know why we have been spared.
In Eurasia, these moths also boom and crash every so often, about as frequently as they do here. So they are behaving pretty much the same where they have existed for a very long time and where they have been present for only a century and a half.
How bad are they?
The moths continue to spread into new parts of the continent. When moths first arrive in an area, their initial outbreak can kill a few vulnerable trees. The percent mortality is usually in the single digits (rarely a true decimation, one out of ten). After that, tree mortality is quite low, but not zero. In 1981 near our house, a large hemlock tree surrounded by oaks was stripped bare and never recovered. In contrast, clusters of hemlocks all survived because they weren’t completely defoliated. None of the oaks and white pines died, even though they lost essentially all their leaves (the pine needles were eaten down to little stubs, but for some reason, not all the way down to the branches).
But even if the trees weren’t killed, were they affected in other ways? Yes. When trees lose all or most of their leaves, their growth slows down. How do we know? By looking at the growth rings of the trees.
OLYMPUS DIGITAL CAMERA
This is a piece of red oak that I cut for firewood in the winter of 1988-89. I know when I cut it because of the distinctly narrow ring from 1981 when the tree was defoliated. In 1981, the tree began to grow in the spring (the coarse cells in the inner portion of the ring) but then grew slowly or not at all until the larvae had finished eating and new leaves could form in the summer. Counting out from 1981, the last ring is 1988 (the one just inside the bark). I cut this tree after the growing season of 1988, but before any growth could occur in 1989.
Tree rings reveal good years and bad years of growth, and insect attack is only one of many possible causes. Shade from other trees produce consistently narrow rings (until the tree casting the shade dies). Drought can slow growth, especially in trees that are rooted in soils that don’t hold a lot of water. Injury to the tree can also be a problem if it is serious. And feeding by many species of insects, indigenous and introduced, can damage trees.
Questions
Has the arrival of Lymantria moths increased the frequency, duration and/or amount of growth reduction in oak trees? Or are moth outbreaks just another cause of growth reduction among those already apparent in the tree ring record?
I tried to answer these questions at six sites in Pennsylvania and New Jersey where the first outbreaks of these moths were well documented by the state forestry departments. I measured the tree rings before and after the first outbreaks, and used observations from foresters, and published records of drought severity, to determine (when possible) the cause of significant declines in growth.
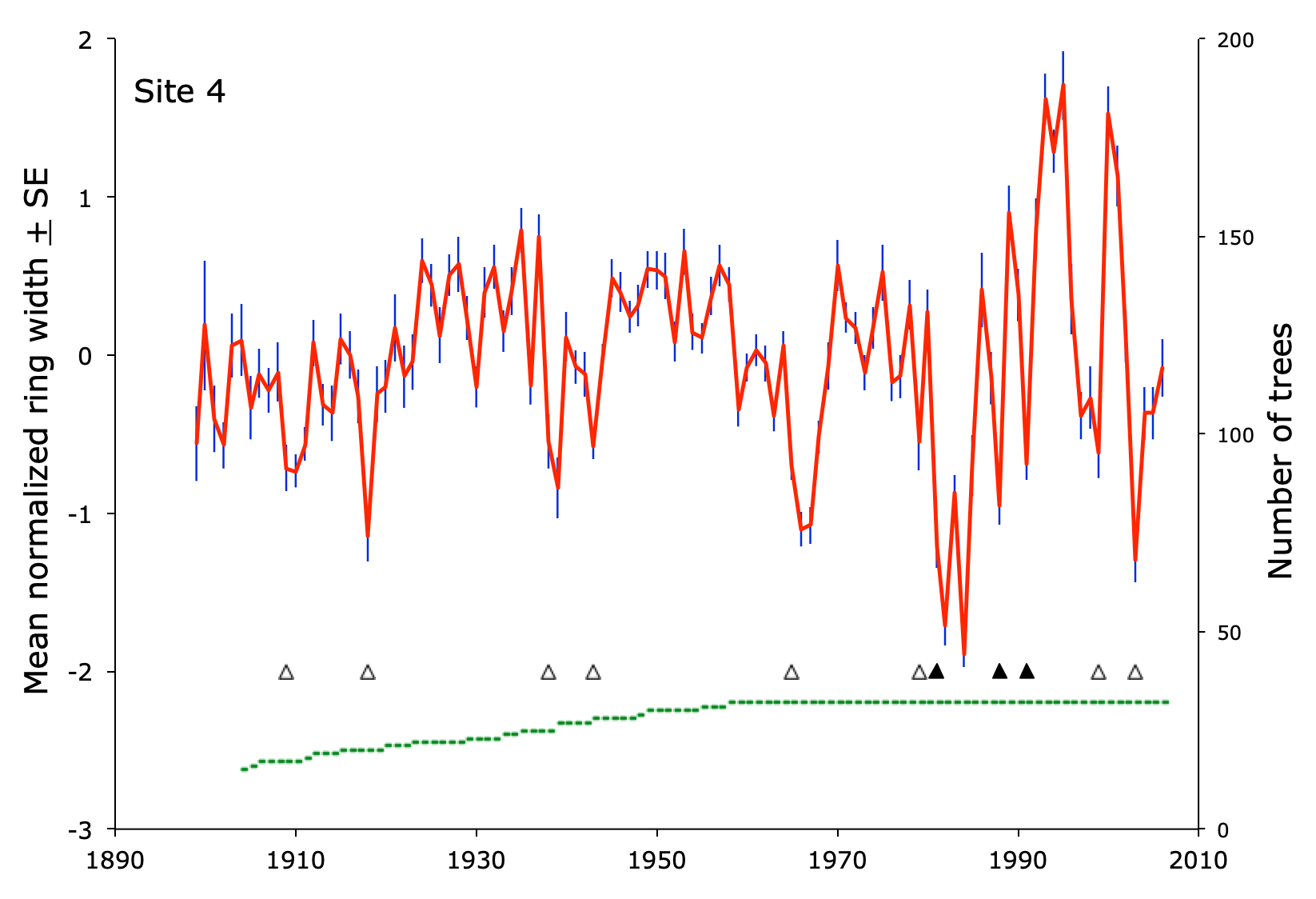
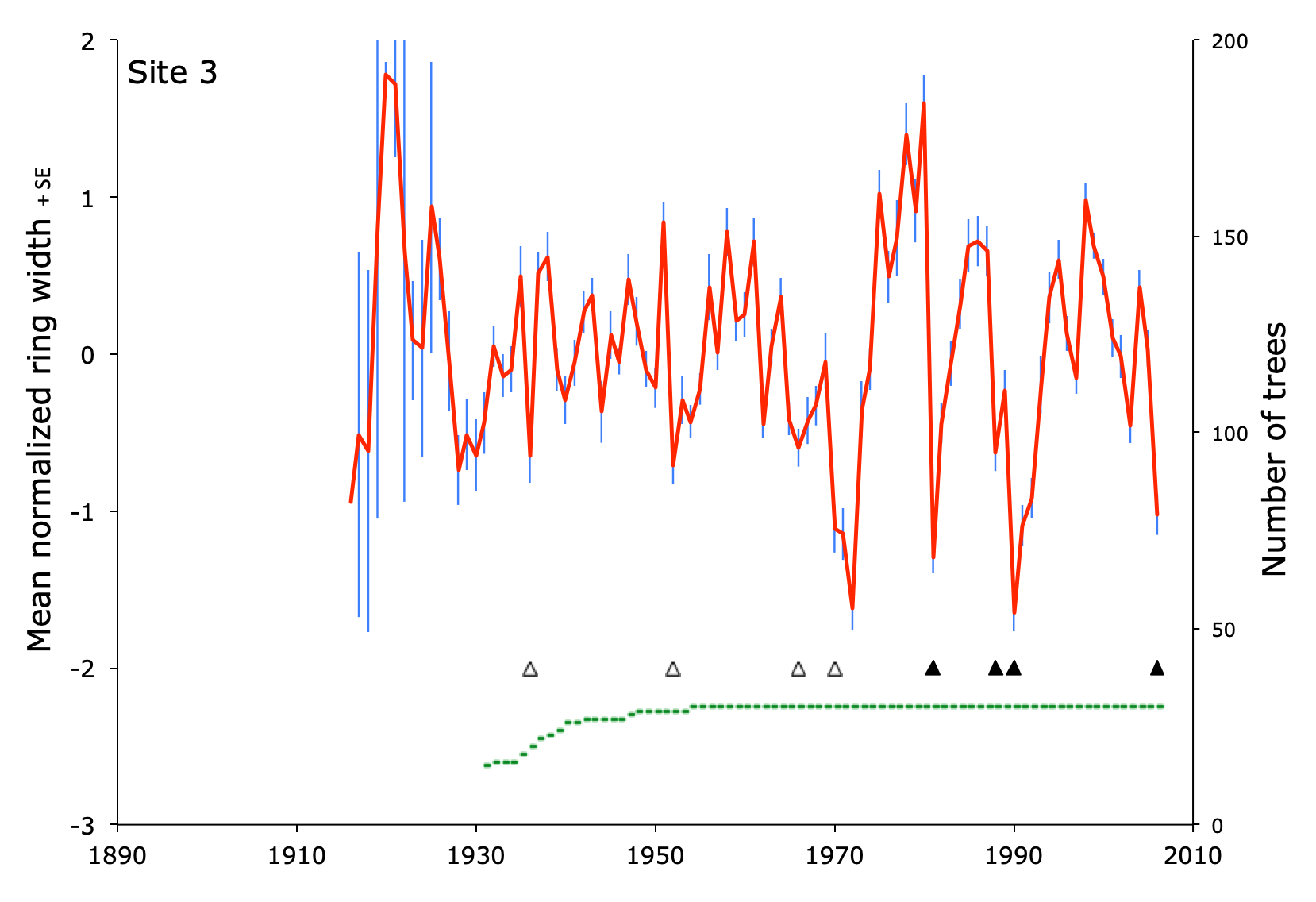
Tree growth, as recorded in the width of tree rings, varies widely over time. Different trees grow at different rates for all kinds of reasons, including genetics, neighbors, soil, weather, and so on. To reduce the individual differences among trees, space, and time, dendroecologists detrend, normalize, and average the results for dozens of trees at a site. The result looks like this graph from site 4 in Pennsylvania.
Years of low growth
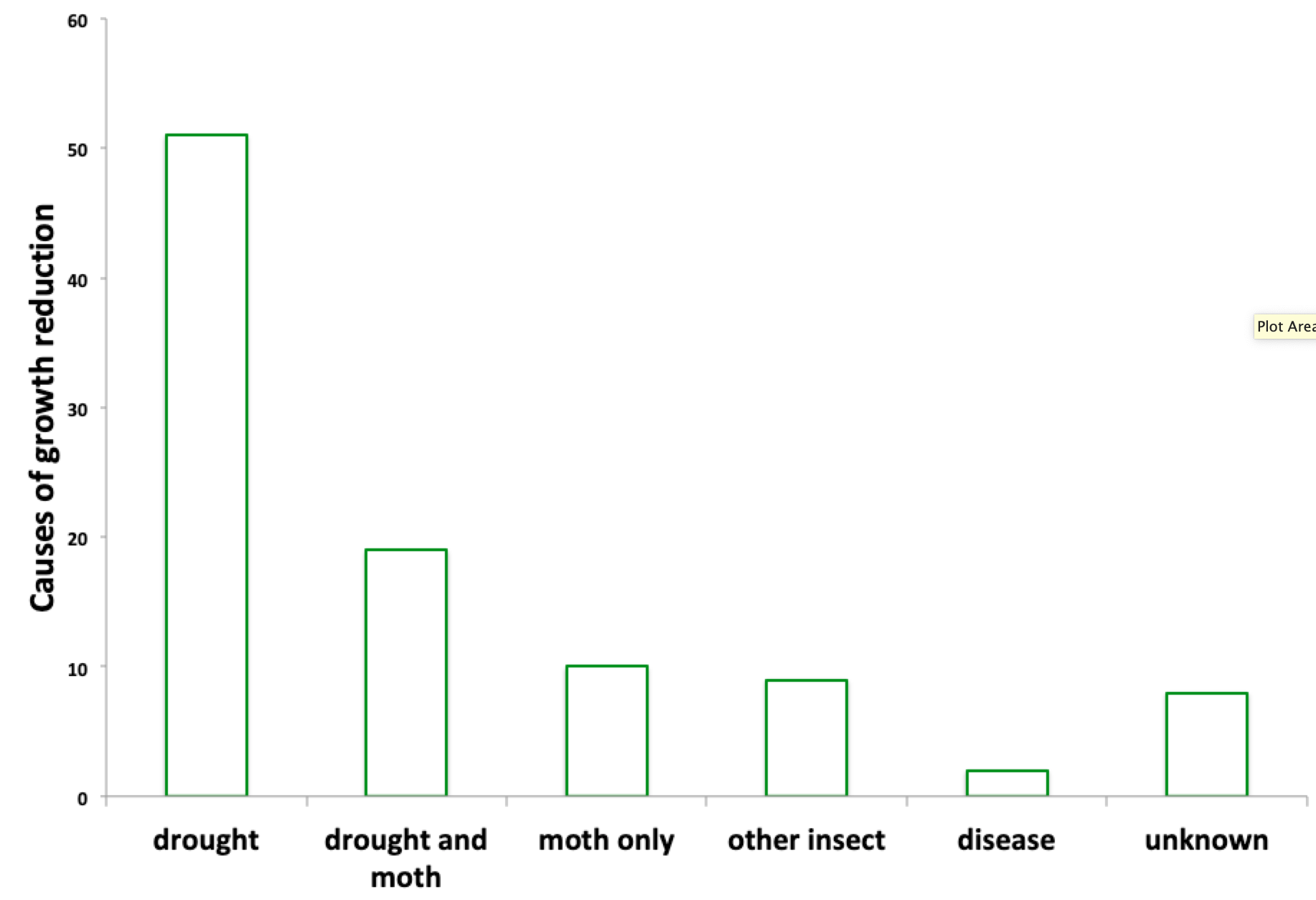
At the six sites, there were 529 site-years with at least fifteen trees present. In 78 of those site-years (15%), growth was reduced by at least one half of a standard deviation.
Fifty-one of the reductions were in drought years, 29 occurred during outbreaks of Lymantria (19 of which were also drought years), nine were associated with outbreaks of indigenous insects (oak leaf tier, oak leaf roller, and scale insects), and two occurred when there was evidence of fungal disease. For eight of the growth reductions, I couldn’t find anything in the drought or forestry records that was associated with low growth.
The magnitude of growth reduction varied widely, but on average, Lymantria outbreaks caused greater reductions in growth than droughts. The mean reduction of growth in drought years was -0.69. When moths were present during droughts, the mean reduction in growth was -0.94. During outbreaks when there was no drought, the mean reduction in growth was -1.20.
But the plot of these results shows something interesting. Growth reductions usually ranged between -0.5 and -1.2 for drought only, drought plus moths, and moths only. In five cases, growth plummeted by more than -1.5. At their worst, Lymantria outbreaks caused the greatest reductions in tree growth over nearly a century. On the other hand, they did so only five times between the time outbreaks began until 2006 (when my data collection ended).
Events of low growth
Reductions in growth often lasted for more than one year at a site. Droughts and insect outbreaks might end after one growing season, or continue for several years. Has the arrival of Lymantria moths changed these events, compared to growth reductions before they arrived?
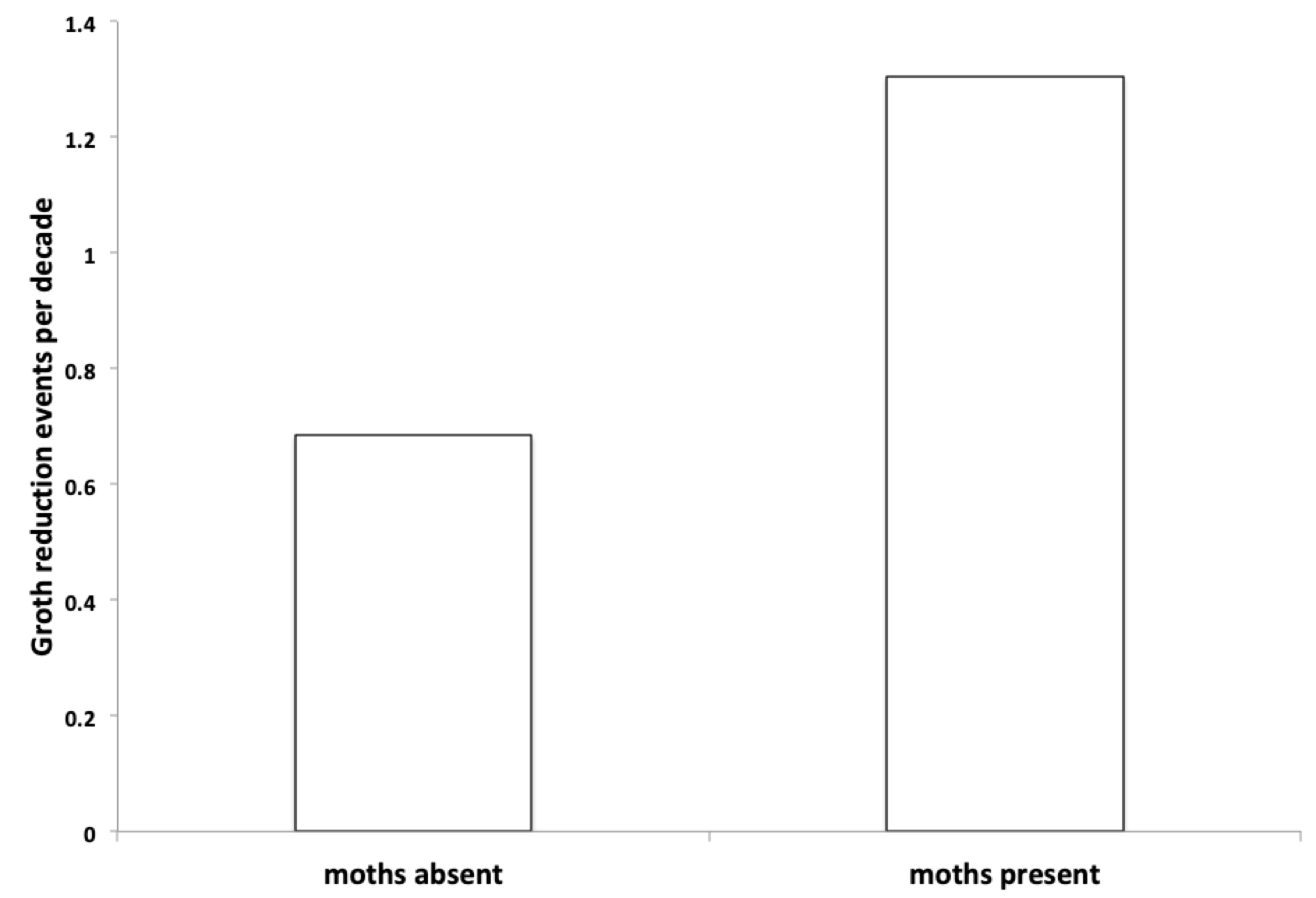
Before there were outbreaks of Lymantria, growth reduction occurred less than once per decade at the six sites. After moth outbreaks began, growth reduction was about twice as frequent, well over one per decade. It is impossible to know why the frequency increased, but this invasive moth almost certainly contributed to the increase.
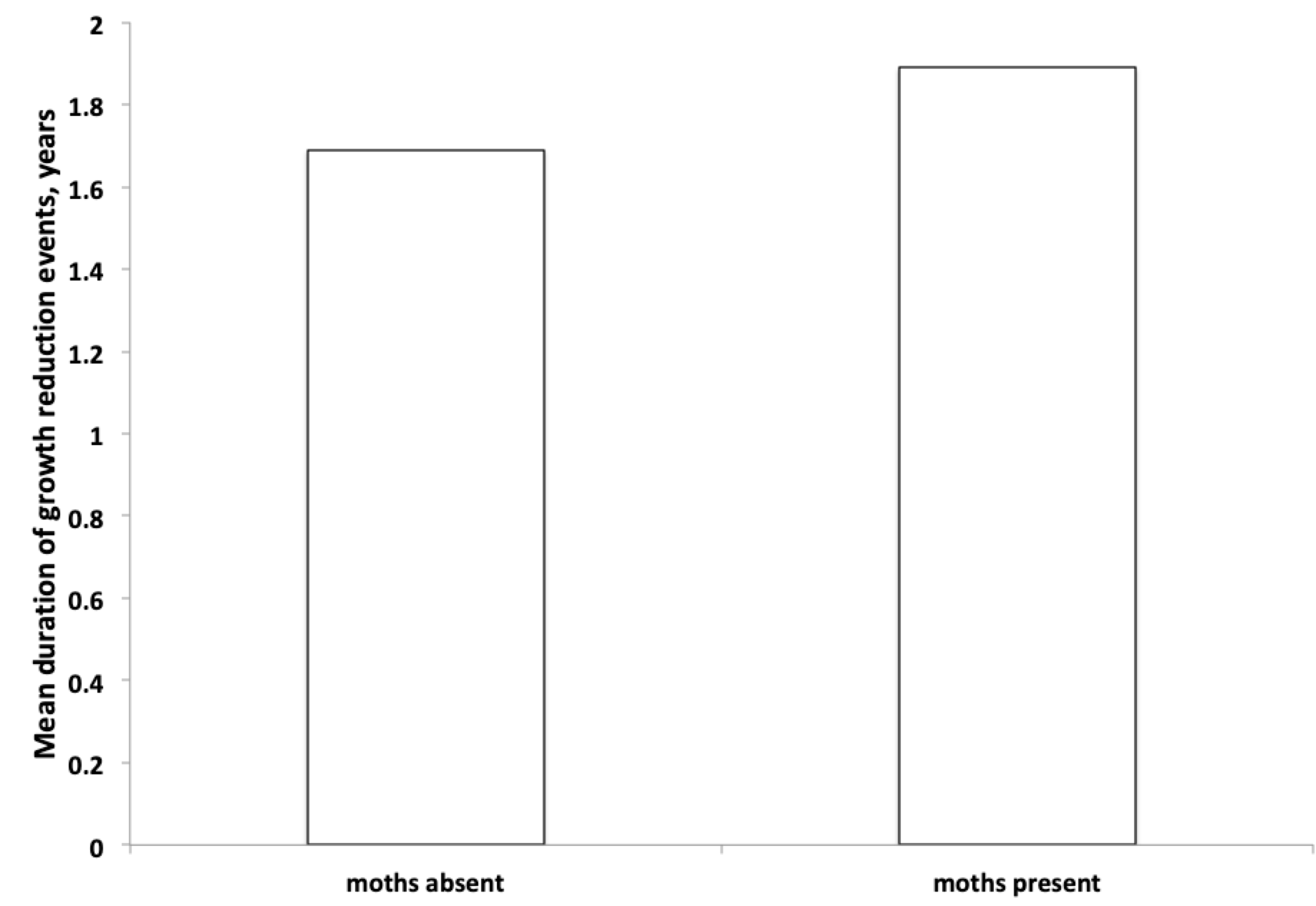
On average, growth reduction events lasted 1.7 years when moths were absent. Duration increased somewhat, to 1.9 years, once moths were present at the study sites. This difference is not statistically significant, and probably not ecologically significant.
The maximum amount of growth reduction during an event increased quite a bit once moths arrived, going from an average of -0.74 down to -1.01. An outbreak of Lymantria moths can be a big deal for tree growth.
But so can an outbreak of the indigenous oak leafroller, Archips semiferanus. At site three, the outbreak began in 1970 and lasted three years, reducing tree ring width to -1.62 in 1972.
After nearly every outbreak of an insect defoliator, whether invasive or indigenous, tree growth quickly shot up to values among the highest seen in the entire tree-ring record. The trees are resilient, for whatever reason, after severe damage from herbivores.
How bad is this invasive insect
There are plenty of invasive pests that are serious problems, none worse than the chestnut blight that wiped out American chestnuts in the twentieth century. Today, the emerald ash borer is killing lots of ash trees, and the hemlock woolly adelgid is killing lots of hemlocks. If we can stop the spread of these introduced pests, we can save a lot of trees.
How serious is Lymantria dispar? Outbreaks of these caterpillars can kill trees, and the survivors have reduced tree growth. Also, the frequency and intensity of growth reduction has increased since the moths arrived.
On the other hand, outbreaks are infrequent, few trees are killed, and the surviving trees seem to bounce back readily after an outbreak. The moths have affected tree growth, but have not radically altered our forests.
Perhaps most significant, there is nothing we can do to stop the spread of this moth. It continues to disperse west, south and north since its release in Massachusetts over a century ago. We will have to tolerate its presence, just as people – and forests – do across Eurasia. We have no choice.
By the way, if you have never heard of the spongy moth, it is because it is the new name for the gypsy moth. The old common name for Lymantria dispar is ethnically insulting, so it has been changed https://www.nytimes.com/2022/03/03/science/spongy-moth-romani.html